Афонин А.А. Метагенофонд симпатрических популяций с
различным уровнем репродуктивной изоляции: на примере ив (Salix L.) / А.А. Афонин, Л.Н. Анищенко, Е.В. Борздыко, Н.В.
Маркелова // Ежегодник НИИ фундаментальных и прикладных исследований за 2010 г.
– Брянск: РИО Брянского государственного университета, 2010. – С. 77–86.
УДК
608.3 + 574.4 + 582.623 + 630*228.7 + 630*165.4
Метагенофонд симпатрических популяций с
различным уровнем репродуктивной изоляции: на примере ив (Salix L.)
А.А. Афонин,
Л.Н. Анищенко, Е.В. Борздыко, Н.В. Маркелова
Брянский государственный университет имени академика
И.Г. Петровского
Предложено понятие «метагенофонд» для обозначения множества популяционных генофондов в составе биоценоза. При высокой вероятности гибридизации метагенофонд рассматривается как единая генетическая система: на примере эколого-генетического комплекса Salix alba – Salix fragilis. При низкой вероятности гибридизации метагенофонд рассматривается как конгломерат популяционных генофондов: на примере ценоза Salix viminalis – Salix dasyclados.
Ключевые слова: ивы, метагенофонд, генетическая безопасность, полиплоидия, гибридизация, репродуктивная изоляция.
Интенсификация селекционного процесса сопровождается
быстрой экспансией новых сортов и пород (включая распространение
ГМ-организмов), что актуализирует проблему генетической безопасности. Выражение
«генетическая безопасность» (genetic safety, genetic security)
используется в самых разных значениях [13]. Чаще всего под генетической
безопасностью понимают состояние
защищенности генетической информации от нежелательных внешних воздействий. В
то же время, генетическая безопасность может рассматриваться как состояние защищенности биологической системы,
определяемая генетической информацией, содержащейся в данной системе. Объединив
указанные подходы, получаем следующее определение: генетическая безопасность – это состояние защищенности генетической
информации, которое определяется самой генетической информацией [3].
Можно
говорить о генетической безопасности биосферы, биомов, экосистем, биоценозов,
популяций и внутрипопуляционных группировок: форм, семей, индивидов (личная
безопасность). В данном случае мы рассматриваем понятие «генетическая
безопасность» применительно к квазинативным экосистемам: внешне похожим на естественные, но сложившимся под влиянием
антропогенных факторов. Квазинативные экосистемы могут складываться стихийно
(вырубки и вторичные леса, заброшенные сельхозугодья, пустыри, полосы отчуждения),
но могут создаваться и целенаправленно (искусственные леса) [12].
Разумеется, степень антропогенного вмешательства в динамику
экосистем изменяется в градиенте «однократное прямое воздействие – постоянное
косвенное воздействие – постоянный максимально возможный контроль» сложным
образом. По характеру влияния антропогенного фактора можно выделить 4 основных
класса квазинативных экосистем:
I. Экосистемы, подвергавшиеся воздействию антропогенного фактора в
прошлом.
II. Экосистемы, постоянно подвергающиеся прямому воздействию
антропогенного фактора в настоящем.
III. Экосистемы, постоянно подвергающиеся косвенному воздействию
антропогенного фактора в настоящем.
IV. Экосистемы, подвергающиеся случайному воздействию антропогенного
фактора в настоящем.
Генетическая безопасность квазинативных экосистем
обеспечивается превышением допустимого (критического) уровня генетического
разнообразия, обеспечивающего внутреннюю и внешнюю устойчивость экосистем, что
минимизирует возможные эволюционные (эколого-генетические) риски.
Существует
два крайних типа генетических рисков:
– загрязнение популяционных генофондов: интеграция в генетическую систему популяций новых
аллелей и изменение частот аллелей (вплоть до элиминации);
– вытеснение генофондов: замещение первоначальных популяционных генофондов
новыми генофондами того же или иного биологического вида.
Живыми
компонентами экосистем являются популяции и формы популяционного ранга,
образующие сообщества (биоценозы). Указанные генетические риски в биоценозе
определяются вероятностью латерального переноса генов (аллелей, гаплотипов).
Генетическая безопасность отдельно взятой популяции определяется её
генетической структурой (генетической системой) и системой воспроизведения
данной популяции. Генетическая безопасность биоценоза определяется его
метагенофондом – множеством популяционных генофондов в составе биоценоза.
Основным
генетическим риском для отдельно взятой амфимиктической квазинативной популяции
является непредсказуемое изменение
генофонда в результате вторжения (инвазии) носителей новой генетической
информации. В данном случае возможно вытеснение аллелей (гаплотипов) дикого
типа новыми аллелями (гаплотипами), повышающими приспособленность их носителей,
но снижающих среднюю приспособленность популяции. Не менее опасная прямая эрозия (вырождение) генофондов (утрата
аллелей и генотипов в результате эффекта генетической воронки). Основным
генетическим риском при ограничениях панмиксии в квазинативных популяциях
(например, при инбридинге, партеногенезе, апомиксисе, вегетативном размножении)
является прямая инвазия иных генотипов,
которые могут обладать более высоким информационно-энергетическим потенциалом и
вытеснять первоначальные генотипы. В результате может происходить замещение
исходных генотипов новыми генотипами с иными свойствами и, как следствие ‒ полное преобразование генетической структуры
популяции. Основным генетическим риском для квазинативных биоценозов является прямая инвазия популяций иноземных видов,
вытесняющих популяции автохтонных видов. В результате происходит
непредсказуемое изменение метагенофонда и переформирование всех экологических
(энергетических) взаимодействий в экосистемах.
Приемлемый
уровень генетической безопасности определяется возможностью сохранения
существующего уровня биоразнообразия на протяжении длительного числа поколений
при отсутствии катастроф. В то же время, оптимальный уровень биоразнообразия
обеспечивает устойчивость экосистем к воздействию внешних факторов. В
результате сохраняется возможность
эволюции сообществ и популяций.
На протяжении последних десятков лет в общественно-научном
сознании сложилось мнение, что основу биоразнообразия составляет
альфа-разнообразие (число видов в экосистеме). Отсюда следует вывод о том, что
монокультура не может быть устойчивой. Однако часто достаточно устойчивыми
являются экосистемы, в которых альфа-разнообразие сравнительно невелико.
Например, в экосистемах равнинных европейских лесов каждый род относительно
крупных организмов, как правило, представлен одним видом (реже двумя или
немногими видами). Изучение популяционно-видовой структуры лесов Сибири (в
меньшей степени подверженных воздействию антропогенных факторов, чем в средней
полосе России) дает возможность утверждать, что высокая устойчивость лесных
экосистем может быть достигнута при низком уровне видового разнообразия
эдификаторов, но высоком уровне их генетического разнообразия [15].
Теоретическое объяснение этого эмпирического факта было предложено нами ранее
[12].
В то же время, известны многовидовые генетически гетерогенные
экосистемы, отличающиеся высокой устойчивостью к воздействию внешних факторов.
Примером таких экосистем служат ивняки, в которых средообразующими
видами-эдификаторами являются ивы (Salix L.) – RS-стратеги, известные высоким
уровнем видового и внутривидового разнообразия. Ивы образуют естественные насаждения (ивняки), входят в состав
разнообразных древесно-кустарниковых сообществ, а также выращиваются в культуре
[16].
Склонность
ив к межвидовой гибридизации известна ботаникам с XVIII в. (Scopoli, 1760; De Candolle,
1832; Kerner, 1860; Wimmer, 1866), а
в XIX в. межвидовые гибриды ив были получены
экспериментально (Wichura, 1854, 1865) [цит. по:
17]. Гибридизация ив широко использовалась в селекционном процессе [1, 18, 19,
21]. Способность ив к межвидовой гибридизации носит дифференциальный характер:
одни виды (даже принадлежащие к разным секциям и подродам) скрещиваются легко,
другие – с трудом. Это дает возможность сравнить характер генетических угроз в
континууме «генетическая система – генетический конгломерат».
В состав
ивовых ценозов средней полосы России входит до 8 видов, включая виды, способные
скрещиваться между собой. Однако фактически мы не наблюдает ни слияния видов,
ни вытеснения, что указывает на существование механизмов, обеспечивающих
эволюционную устойчивость ивовых сообществ [8, 9].
Для
выявления возможных механизмов эволюционной устойчивости генофондов популяций и
метагенофондов сообществ нами были выбраны массовые виды ив: белая (S. alba) и её гибриды с ивой ломкой (S. fragilis), корзиночная (S. viminalis) и
шерстистопобеговая (S. dasyclados).
Эти виды отличаются друг от друга уровнем плоидности (геномными числами) [27,
29], характеризуются разными географическими и эдафо-фитоценотическими
ареалами.
Ива
белая (Salix alba L.) и её
гибриды с ивой ломкой (Salix fragilis L.)
Ива белая
– крупное дерево первой величины с прямым толстым стволом. Листья
продолговато-эллиптические, ланцетовидные, иногда очень узкие, длиной 70...100
(40...130) мм при ширине 16...22 (11...27) мм. Опушение шелковистое: волоски
серебристые, длинные, прижатые, вытянутые параллельно средней жилке.
Прилистники ланцетовидные, мелкие (не более 5 мм), рано опадающие.
Тетраплоид:
2n=4х=76. Длина пыльцевых зерен 30,0±0,08 мкм
[7].
Ареал
евро-азиатского типа, бореально-средиземноморского подтипа: Средняя и
Атлантическая Европа, Западная Европа (кроме Скандинавии), Средиземноморье,
Кавказ, Средняя и Малая Азия, Иран, Западная, Юго-Западная, Юго-Западная и
Восточная Сибирь, Гималаи, Китай. В России северная граница ареала проходит по
линии Новгород – Вологда – Киров – Пермь, в Западной Сибири доходит до р.
Енисея.
Типичный
пойменно-аллювиальный вид. Приурочена к хорошо дренированным почвам с
постоянным увлажнением. Рассеянно в поймах всех рек, за исключением самых
малых. Образует сплошные насаждения (ленточные ивняки) вдоль рек и озер,
тяготеет к прирусловой части поймы. В центральной пойме и вне пойм – в виде
солитеров (одиночных особей). Цветет относительно поздно (конец апреля – начало
мая).
Внутривидовые
формы: f. pendula Host (L.) (плакучая крона), f.vitellina
pendula Rend. (плакучая крона с блестяще-желтыми побегами), f. populiforme
Leveiell = f. piramidalis Host (пирамидальная крона), f. vitellina Stokes = S. vitellina L.
(блестяще-желтые зрелые однолетние побеги), f. britzensis
Spaeth. (красные или краснеющие побеги), f. splendens Bray = var. argentea
Wimm. = var. regalis Anderss. (серебристо-блестящие снизу листья), f. coerulea Wimm. = S. coerulea Smith
(голубоватые листья), f. ovalis Wimm. (продолговато-эллиптические
листья с округлым основанием и слабо заостренной верхушкой; ширина пластинки
достигает максимума в средней части).
Вследствие
географической изменчивости выделяется подвид S. alba ssp. micans Anderss.
и разновидность S. alba var. australior Anderss. Близкий вид: S.excelsa S.G. Gmelin – ива
южная (каратал). В естественном состоянии на территории средней полосе России
эти таксоны не встречаются.
Ива
ломкая (S. fragilis L.; син. S.
decipiens Hoffman
1791) традиционно считается самостоятельным видом малоазиатского и
закавказского происхождения, проникшим в Европу несколько сот лет назад. Как и
ива белая, это тетраплоид: 2n=76. Длина
пыльцевых зерен 29,4±0,10 мкм [7].
Используется
так же, как и ива белая, но в меньшей степени. Культивируется клон с густой
шарообразной кроной (S. fragilis var. sphaerica Hryniew.
= S. fragilis L. f. bullata hort.).
Однако
фактически этот вид изучен недостаточно. По мнению И.В. Беляевой [22], все
европейские особи ивы ломкой представляют собой гибриды S. alba и S. euxina I.V. Belyaeva.
Последний вид, действительно, произрастает на территории Северной Турции и
прилегающего Кавказа. Таким образом, фактически на территории средней полосы
России мы имеем дело с комплексом [S. alba – S. ´ fragilis
– S. euxina], в
котором осуществляется латеральный перенос генов.
Ива
корзиночная (Salix viminalis L.)
Быстрорастущий высокий кустарник. Листья очень узкие,
часто почти линейные, иногда саблевидно изогнутые. Длина 80...125 (45...160)
мм, при ширине 10...15 (7...18) мм. Прилистники мелкие, ланцетовидные, рано
опадающие.
Диплоид: 2n=2х=38.
Длина пыльцевых зерен 29,0±0,12 мкм [7].
Ареал
евроазиатского бореального типа: Средняя и Атлантическая Европа (на севере
европейской части России доходит до Белого моря), Западная и Восточная Сибирь,
Монголия, Китай, Индостан.
Типичный
пойменно-аллювиальный вид, где образует сплошные насаждения вдоль рек и ручьев
по аллювиальным наносам, на песчаных отмелях, обрывах. Избегает избыточного застойного
увлажнения. Вне пойм редко, иногда вдоль дорог, в нарушенных местообитаниях.
Цветет рано (II половина апреля – первые числа
мая). За рубежом выращивается на энергетических плантациях для получения
биомассы [24, 26].
Известна
под синонимами: S. serotina Pallas,
1776, S. gmelinii Pallas,
1788, S. rossica Nas.
1936, S. pseudolinearis Nas.
1936, S. verviminalis Nas.
1936, S. semiviminalis E. Wolf, S. strobilacea Nas.
Внутривидовые формы: – f. culta hort.
(культивируется). Подвиды: S. viminalis ssp. rossica Nas. –
(отличается более широкими листьями (узко ланцетовидными) с округленным
основанием, туповатыми почками и более поздним цветением); S.viminalis ssp. schwerinii E. Wolf (ива Шверина, Дальний Восток).
Ива
шерстистопобеговая – Salix dasyclados Wimm.
Крупный
быстрорастущий кустарник, реже дерево. Порослевые побеги прутьевидные, очень
мощные. Листья крупные, широко ланцетовидные, длиной 70...105 (40...140) мм при
ширине 20...30 (15...35) мм. Прилистники очень крупные (до 10 мм и более),
серповидные; долго сохраняющиеся.
Гексаплоид:
2n=6х=114. Длина пыльцевых зерен 33,1±0,08 мкм
[7].
Ареал
евроазиатского бореального типа (но более продвинута к северу, чем
корзиночная): Европа, Сибирь, Дальний Восток, Монголия, Китай, Япония. В
европейской части России южная граница проходит в Верхне-Днепровском районе, на
севере доходит до Белого моря.
На сырых
почвах, в понижениях центральной и прирусловой поймы, вне поймы обычна на сырых
лугах, вдоль дорог; редко в посадках. Отсутствует на болотах с избыточно застойным
увлажнением. Цветет очень рано, сразу после установления положительных ночных
температур.
Синонимы: S. burjatica
Nas., S. jacutica Nas. Однако
существует точка зрения [23], согласно которой иву шерстистопобеговую следует
называть Salix gmelinii Pall. (1789) – ивой Гмелина. Названия Salix dasyclados Wimm., S. burjatica Nas., S. jacutica Nas. в этом
случае становятся избыточными. Не исключено [20], что данный вид имеет
гибридное происхождение.
Степень
репродуктивной изоляции (способность к межвидовой гибридизации) перечисленных
видов различна (табл.) [4, 6].
Таблица
– Распространенные межвидовые гибриды ив с участием S. alba, S. fragilis, S. viminalis, S. dasyclados
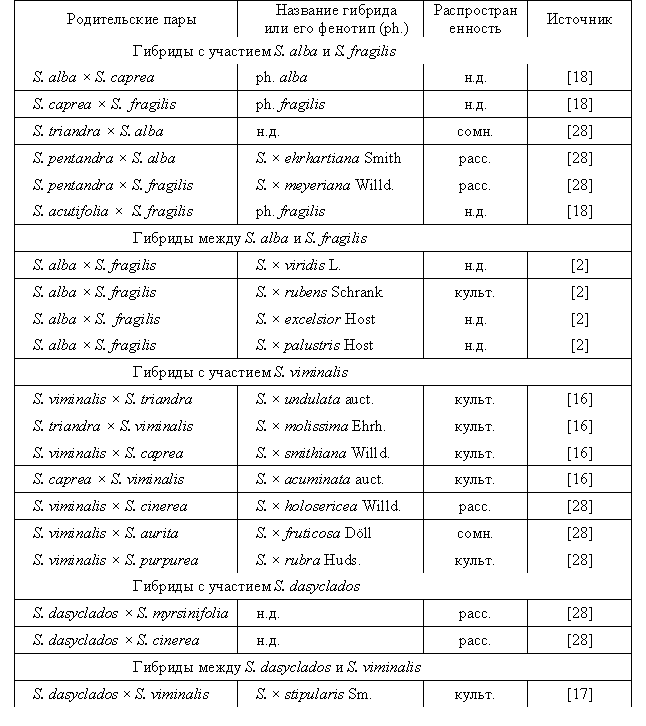
Примечание: культ. – культивируется, расс. – рассеянно в природе, сомн. – существование гибрида сомнительно, н.д. – нет данных
Материал для исследования собирался в
демонстрационно-экспериментальном салицетуме Брянского государственного
университета и в квазинативных ивняках Брянского лесного массива (в зеленой
зоне г. Брянска).
Для создания салицетума
использовались семьи и контрольные популяции. Каждая семья представляет собой
семенное потомство, полученное от одной особи. В отдельно взятой семье снижен
уровень генетического разнообразия. В то же время, сеянцы (особи нулевого
возраста) еще не успевают пройти через сито естественного отбора, что позволяет
обнаружить редкие варианты признаков, не встречающиеся в группах поздних
возрастов (от виргинильных до сенильных стадий онтогенеза). При семенном
размножении путем инцухта (включая беккроссы) возможно дополнительное
выщепление рецессивных признаков. Каждая контрольная популяция представляет
собой совокупность саженцев, выращенных из черенков, срезанных с разных особей.
При должной репрезентативности выборки уровень генетического разнообразия
достаточно высокий. Однако обнаружение редких вариантов признаков в этом случае
менее вероятно, чем в семьях.
При сборе материала в
квазинативных ивняках достигается максимально высокий уровень генетического
разнообразия. Однако (как и в случае контрольных популяций) обнаружение редких
вариантов признаков в этом случае менее вероятно, чем в семьях. Кроме того,
материал является абсолютно разновозрастным, что вносит дополнительный шум в
виде возрастной изменчивости.
Метагенофонд комплекса S. alba – S. fragilis
Для изучения метагенофонда комплекса S. alba – S. fragilis
использовались семьи, полученные посевом семян, собранных в квазинативных ивняках
с различным характером воздействия антропогенного фактора.
Семья al03-02-хх выращена из семян, собранных 14.06.2003 г. с особи,
обладающей типичными признаками вида. Место сбора: центральная пойма р. Десны,
ассоциация Salicetum albae, в черте г. Брянска, в
точке с координатами 53,2208 с.ш., 34,38009 в.д. Это сообщество в наибольшей степени
подвержено влиянию антропогенного фактора (все классы: I…IV). Здесь
наряду с типичными S. alba часто
встречаются гибриды S. alba
´ S. fragilis. Типичные особи S. fragilis (S. decipiens) с абсолютно неопушенными листьями не
обнаружены. В настоящее время в
салицетуме сохранено три высокопродуктивных устойчивых ортета: al03-02-05♂, al03-02-08♀, al03-02-10♂.
Для закладки семьи al03-06-хх
использовались семена, собранные 15.06.2003 г. с особи, обладающей типичными
признаками вида. Место сбора: прирусловая пойма р. Десны, ассоциация Salicetum albae, за чертой г. Брянска, в точке с координатами
53,189762 с.ш., 34,268689 в.д. Данное сообщество в
наименьшей степени подвержено влиянию антропогенного фактора (класс IV). Здесь не отмечены гибриды S. alba ´ S. fragilis, но
обнаружена S. alba f. ovalis. В настоящее время в салицетуме сохраняется ортет al03-06-01♂.
Для получения синтетической семьи S. alba второго порядка (F1, 2008 г.) ортет al03-02-08♀ был опылен
пыльцой al03-06-01♂. В результате
была получена синтетическая семья al08-(28×61)-хх.
Гибридная семья al´fr03-02хх получена из семян, собранных 14.06.2003 г. с особи, обладающей типичными признаками
гибрида S. alba ´ S. fragilis. Место сбора:
центральная пойма р. Десны, ассоциация Salicetum fragilis, в
черте г. Брянска, в точке с координатами 53,255248 с.ш., 34,382311 в.д. Это сообщество в наибольшей
степени подвержено влиянию антропогенного фактора (все классы: I…IV). Все сеянцы
обладали типичными чертами гибридов S. alba
´ S. fragilis. В настоящее время в салицетуме сохраняется ортет al´fr03-02-06♀, от которого не
удалось получить полноценные семена.
Гибридная семья vp94-01-хх получена
из семян, собранных 7.06.1994 г. с культивара ивы плакучей (cv ′vitellina pendula′)
неизвестного происхождения. Место сбора: частично окультуренный пустырь в черте
г. Брянска, в точке с координатами 53,255851 с.ш.,
34,372158 в.д. Это местообитание в
наибольшей степени подвержено влиянию антропогенного фактора (классы I…IV). Среди
сеянцев F0
наблюдалось расщепление по форме кроны, морфологии листовых пластинок и
прилистников, окраске годичных побегов. Для получения последующего гибридного
поколения (F1)
использовались пестичные ортеты vp94-01-01♀
(желтопобеговая), vp94-01-04♀
(краснопобеговая) и тычиночный ортет vp94-01-02♂
(зеленопобеговый). Среди полученных сеянцев vp99-01-01-хх
также наблюдалось расщепление по форме кроны, морфологии листовых пластинок и
прилистников, окраске годичных побегов. В дальнейшем (2006 г.) были получены
сеянцы от скрещивания высокопродуктивного желтопобегового ортета vp99-01-01-01♂ с краснопобеговыми ортетами vp 99-01-01-02♀ и vp
99-01-01-03♀.
Для получения синтетических гибридных семей второго
порядка (2006 г.) использовались «возвратные» скрещивания al03-02-08♀ ´ vp99-01-01-01♂. Гибридные сеянцы обладали чертами гибридов S. alba ´ S. fragilis,
не отличаясь какими-либо особыми качествами.
В 2010 г. трехлетние сеянцы синтетической семьи al08-(28×61)-хх), сохранив
типичные черты S. alba, проявили расщепление по длине
лидерных побегов. Отчетливы выделились три размерных класса: 60…65 см, 95…105
см, 150…170 см. Отношения средних приростов в смежных классах составили
1,6…1,7. Кроме того, один сеянец дал прирост в 210 см. В 2005 г. среди
трехлетних сеянцев всех древовидных ив посева 2003 г. (al03-02-хх, al03-06-хх, al´fr03-02хх, vp94-01-хх) было обнаружено
сходное расщепление: 50…65 см, 80…120 см, 140…180 см. Прирост лидерного побега
материнской особи al03-02-08♀
составил 150 см, а лидерного побега отцовской особи al03-06-01♂ – 180 см [5]. Таким образом, в потомстве
двух высокопродуктивных особей (al03-02-08♀
и al03-06-01♂) обнаружено
примерно такое же расщепление по приросту лидерных побегов, как и в выборках из
разных популяций ивы белой и ее гибридов. На этом основании можно утверждать,
что высокая продуктивность ивы белой может быть обусловлена гетерозисом. Это
обстоятельство необходимо учитывать в семеноводстве.
Метагенофонд комплекса S. viminalis – S. dasyclados
Для закладки семьи vi01-01-хх
S. viminalis использовались семена, собранные 20.05.2001 г. с особей, обладающих
типичными признаками вида. Место сбора: понижение прирусловой поймы р. Десны,
ассоциация Salicetum albae, в черте
г. Брянска, в точке с координатами 53,224202 с.ш., 34,366343 в.д. Данное
сообщество в незначительной степени подвержен влиянию антропогенного фактора
(классы I и IV). Весной
2004 г. были собраны и высеяны семена с особи vi01-01-02♀,
опыленной пыльцой высокопродуктивной особи vi01-01-05♂. Большинство полученных сеянцев F1 (vi04-01-02-01…11) сохранили типичные признаки вида, но один
сильный сеянец vi04-01-02-11♀ резко отличался от типа S. viminalis широкими короткими листьями и широкими прилистниками. Получить
потомство от данного сеянца не удалось. Весной 2007 г. были собраны и высеяны
семена с особей vi04-02-02♀, vi04-02-04♀, vi04-02-10♀, опыленных пыльцой рамет vi01-01-05♂.
Все сеянцы F2/b обладали типичными видовыми признаками S.viminalis. К настоящему времени из всего посева 2007 г. в
салицетуме сохранилась ветвь 07-02-02-хх (три сеянца). Весной 2009 г. были
собраны и высеяны семена с раметы vi04-02-04♀, опыленной пыльцой раметы vi01-01-05♂.
Черенки для создания контрольной популяции vi09-1 были срезаны
22.09.2009 г. с особей, обладающих типичными признаками вида. Место сбора:
прирусловая пойма р. Десны, ассоциация Salicetum albae, в черте г. Брянска, в точке с
координатами 53,292023 с.ш., 34,307586
в.д. Хранились при tº=10
ºС. Высажены в салицетум весной 2010 г.
Семья da01-01-хх
S. dasyclados получена из семян, собранных 20.05.2001 г. в центральной пойме р. Десны, в пределах
ассоциации Salicetum triandro-viminalis, с центром в точке с координатами 53, 220569 с.ш.,
34,371874 в.д. Все сеянцы обладали типичными видовыми признаками S.dasyclados. Из полученной семьи da01-01-хх был сохранен
один сеянец (da01-01-07♀) с высокой продуктивностью и
относительной устойчивостью к грибковым заболеваниям. Весной 2009 г. одна из
рамет клона da01-01-07♀ была опылена пыльцой
высокопродуктивного клона ивы корзиночной (vi01-01-05): годичный
прирост лидерного побега в пятилетнем возрасте составил 375 см. Все сеянцы,
выращенные из семян, полученных в результате этого опыления, в 2009-2010 гг.
обладали типичными признаками ивы шерстистопобеговой.
Семена
для создания семьи da10-01-хх были собраны
15.05.2010 г. с особи, обладающей типичными признаками вида. Место сбора:
прирусловая пойма р. Десны, ассоциация Salicetum albae, в черте г. Брянска, в точке с координатами
53,292023 с.ш., 34,307586 в.д. (удалена
от точки сбора семян семьи da01-01-хх на 8,9 км). В течение 2010 г. сеянцы da10-01-хх выращивались
в контейнерах с торфо-минеральным грунтом.
Черенки
для создания контрольной популяции da10-2-хх были срезаны 7.05.2010 г. с особей, обладающих
типичными признаками вида. Место сбора: понижение центральной поймы р. Десны,
ассоциация Salicetum triandro-viminalis, в черте г. Брянска, в точке с координатами 53, 220569 с.ш., 34,371874 в.д.
(там же, где семена для создания семьи da01-01-хх).
Большинство
полученных сеянцев S. viminalis F2/b (vi09-04-01…08) сохранили типичные признаки вида, но два
сеянца резко отличаются от типа S.
viminalis широкими короткими листьями и широкими прилистниками.
Для сравнения использовались листья верхней формации (5-й лист от вершины
побега). Ширина листовой пластинки составила 8…12 мм (в среднем 9,0 мм), длина
– 95…125 мм (в среднем 111,3 мм), индекс продолговатости – 9,2…13,9 (в среднем
11,6). Однако два сеянца резко отличаются от типа S. viminalis широкими листьями – 24…26 мм – при нормальной длине (120 мм). Прилистники у этих
сеянцев также относительно широкие (узко серповидные). Формы ивы корзиночной с
широкими листьями мы предлагаем обозначать как широколистные (Salix viminalis f. latifolia), а сами листья линейно-продолговатыми (lineare-oblongum).
Все
сеянцы S. dasyclados 2009
г., полученные от раметы da01-01-07♀, опыленной пыльцой раметы vi01-01-05♂, сохранили
типичные признаки вида. Более того, внешне они оказались более однородными, чем
черенковые саженцы из контрольной популяции. Это обстоятельство указывает на
возможность апомиктического происхождения семян данного посева.
Сеянцы
посева 2010 г. (семья семьи da10-01-хх) в конце вегетационного
периода разделись на две группы: низкорослые
(высотой 50…90 мм) и высокорослых (h=160…405
мм).
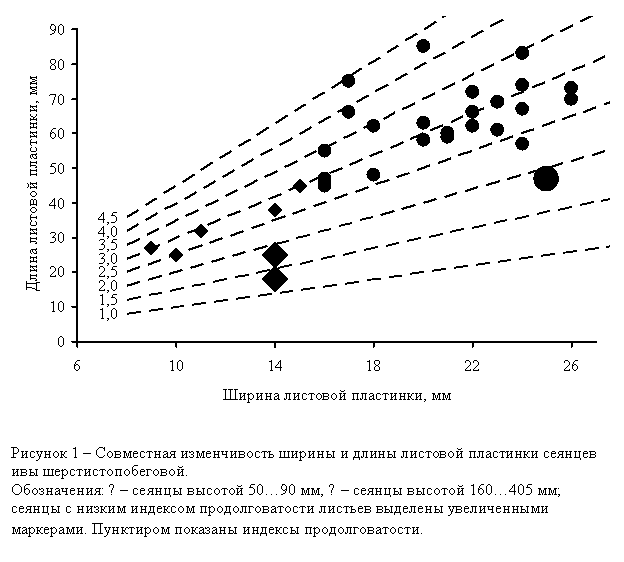
Анализ совместной
изменчивости ширины и длины листовой пластинки (рис. 1) показал, что и среди
низкорослых, и среди высокорослых сеянцев данной семьи S. dasyclados, наряду с типичными особями, встречаются формы с укороченными
листовыми пластинками (Y/X<2,0, вплоть до Y/X<1,3),
При этом уменьшение длины листовых пластинок несколько компенсируется
увеличением их ширины. Такие формы мы предлагаем обозначать как коротколистные
(Salix dasyclados f. brevifolia), а тип конфигурации листовых пластинок – как широко
эллиптические (late ellipticum). С учетом возможного изменения номенклатуры данного
таксона в дальнейшем данная форма может получить название Salix gmelinii f. brevifolia. В природных популяциях подобные особи не обнаружены.
Однако сама возможность их существования свидетельствует в пользу гипотезы о
гетерозисной природе, по крайней мере, некоторых высокопродуктивных особей S. dasyclados. Кроме того, выявленные формы представляют интерес как иллюстрация
генетической гетерогенности природных популяций, а именно – появление новых
вариантов в потомстве типичной особи. Выращивание и дальнейшее семенное
размножение обнаруженных коротколистных особей позволит использовать их как для
изучения закономерностей наследования количественных признаков древесных
растений, так и в селекции S. dasyclados на разнообразие и отличимость [10, 11].
Для сравнения параметров листьев одновозрастных сеянцев
ив корзиночной и шерстистопобеговой была построена совместная диаграмма
рассеяния (рис. 2).
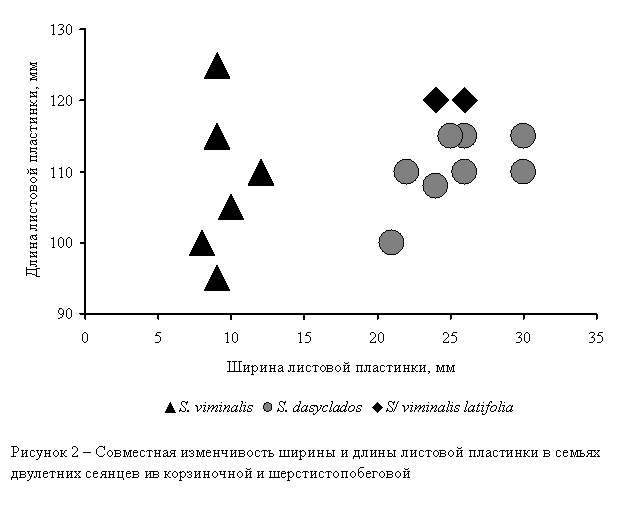
Сравнение параметров двух рассматриваемых выборок
показало, что в данном случае различие индексов продолговатости обусловлено
различиями в ширине листовых пластинок. Учитывая результаты, полученные по
семье da10-01-хх, мы приходим к
следующему выводу: если у ивы шерстистопобеговой изменение конфигурации
листовых пластинок обусловлено, в первую очередь, их укорочением, то у ивы
корзиночной – увеличением их ширины при сохранении длины.
Таким образом, несмотря на определенный параллелизм в
морфогенезе листьев, следует признать, что генетические основы морфогенеза у
этих видов различны: широколистность у S. viminalis
возникает за счет увеличения ширины листовой пластинки без изменения её длины,
а коротколистность у аберрантных возникает за счет уменьшения длины листовой
пластинки без существенного изменения её ширины.
Сравнительный анализ метагенофондов
комплексов S. alba – S. fragilis и S. viminalis – S. dasyclados
Метагенофонд симпатрических популяции S. alba и S. fragilis
образует единую генетическую систему. Гибриды S. alba
´ S. fragilis оказываются вполне плодовитыми при возвратных
скрещиваниях. Спонтанной гибридизации этих видов способствует не только
филогенетическое родство (оба вида принадлежат
к секции Salix Dum. 1825
(Albae)), но и одинаковый уровень плоидности (2n=4х=76), и одновременное цветение. Однако
эколого-генетический комплекс S. alba – S. fragilis обнаруживается
только в составе квазинативных экосистем, подвергающихся сильному
антропогенному воздействию. В этом случае возможен латеральный перенос генов от
S. alba к S. fragilis и
обратно. В экосистемах, менее подверженных антропогенному влиянию, сохраняется
только чистый вид S. alba,
представленный как типичными, так и редкими формами. J. Čhmelar
und W. Meusel [25] считают, что гибриды оказываются менее
конкурентоспособными, чем «чистые» виды, и поэтому не могут встраиваться в
ценозы, близкие к естественным. Таким образом, реальная угроза генетического
загрязнения более вероятна в культурах и антропогенных ландшафтах.
Метагенофонд
симпатрических популяции S. viminalis и S. dasyclados единую генетическую систему не образует, и может
рассматриваться как конгломерат популяционных генофондов двух этих видов. Несмотря на признаваемое филогенетическое родство S. viminalis и S. dasyclados (оба вида принадлежат к секции Vimen Dum. (Viminella Ser.)) и способность образовывать гибриды,
фактического латерального переноса генов не возникает. Этому препятствуют как
разные уровни плоидности (2n=2х=38
и 2n=6х=114),
так и разорванность в периоде цветения [16]. В этом случае генетическое
загрязнение маловероятно, но возникает угроза вытеснения: фактически на хорошо
аэрируемых, но малоплодородных почвах доминирует S. viminalis, а на переувлажненных, но более
плодородных почвах – S. dasyclados, и только при
перекрывании эдафо-фитоценотических ареалов и в нарушенных местообитаниях оба
вида встречаются совместно [14].
Экстраполируя полученные результаты на другие таксоны,
можно реально оценить популяционно-генетические риски. Полученные знания можно
рассматривать как вклад в разработку доктрины генетической безопасности
сообществ и экосистем.
Список литературы
1.
Альбенский А.В. Селекция
древесных пород и семеноводство. – М.: Гослесбумиздат, 1959. – 306 с.
2.
Анциферов Г.И. Ива.
1984. – М.: Лесная промышленность, 1984. – 101 с.
3.
Афонин А.А. Генетическая
безопасность квазинативных экосистем / А.А. Афонин, Н.В. Маркелова //
Экологическая безопасность региона: сб. ст. III Междунар. научно-практ. конфер. ест.-геогр. ф-та
(Россия, г. Брянск, 21–22 октября 2010 г.). – Брянск, Изд-во «Курсив», 2010. –
С. 56–59.
4.
Афонин А.А. Гибридные ивы
Брянского лесного массива // Лесоведение, экология и биоресурсы: Матер.
междунар. научно-производств. конф. «Брянщина – родина отечественного и
мирового высшего лесного образования». – Брянск: БГИТА, 2005. – С. 143–146.
5.
Афонин А.А. Динамика
сезонного прироста по диаметру сеянцев древовидных ив / А.А. Афонин, Н.А.
Лукутина // Вестник БГУ: Естественные и точные науки. – 2007. – № 4. – Брянск:
РИО БГУ, 2007. – С. 115–118.
6.
Афонин А.А.
Дифференциальная гибридизация ив Брянского лесного массива // Лесоведение,
экология и биоресурсы: Матер. междунар. научно-производств. конфер. «Брянщина –
родина отечественного и мирового высшего лесного образования». – Брянск: БГИТА,
2005. – С. 141–143.
7.
Афонин А.А. Изменчивость
длины пыльцевых зерен аборигенных видов ив Брянского лесного массива // ИВУЗ.
Лесн. журн. – 2006. – № 5. – С. 25–34.
8.
Афонин А.А. Механизмы
эволюционной устойчивости симпатрических видов ив // Современные технологии
воспроизводства целевых лесов в Юго-Западном регионе Центральной части России:
Материалы междунар. науч.-производств. конф. – Брянск: Брянская гос.
инж.-технол. академия, 2002. – С. 128–136.
9.
Афонин А.А.
Моделирование устойчивости лесных сообществ в условиях конкуренции между
близкими видами // Химико-лесной комплекс – проблемы и решения: Всеросс.
науч.-практ. конф. – Т. 1. – Красноярск: Сибирский гос. техн. ун-т, 2002. – С.
34–37.
10. Афонин А.А. Морфодинамика однолетних побегов ивы
шерстистопобеговой и перспективы использования концепции морфодинамических
аттракторов для оценки генетического потенциала природных популяций / А.А.
Афонин, Л.Н. Анищенко // Стратегия устойчивого развития регионов России: Сб.
материалов III Всеросс. научно-практ. конфер.
– Новосибирск: Изд-во НГТУ, 2010. – С. 125–132.
11. Афонин А.А. Эколого-генетический потенциал природных
популяций ивы шерстистопобеговой / А.А. Афонин, Е.В. Борздыко, Л.Н. Анищенко //
Фундаментальные науки и практика: Сб. научн. трудов 3-й Международной
телеконференции (Томск, 25 октября – 6 ноября, 2010).
– Томск: СибГМУ, 2010. – С. 159–160.
12. Афонин А.А. Сохранение и приумножение уровня
биологического разнообразия в квазинативных экосистемах / А.А. Афонин, Л.И.
Булавинцева // Биологические эффекты малых доз ионизирующей радиации и
радиоактивное загрязнение среды: Материалы Междунар. конф. – Сыктывкар: Коми НЦ
УрО РАН, 2009. – С. 391–393.
13. Баранов А.С. Популяционно-экологические риски при
выращивании генетически модифицированных организмов (ГМО) // Современное
состояние и пути развития популяционной биологии: Материалы X Всеросс. популяционного семинара (г. Ижевск, 17-22
ноября 2008 г.). – Ижевск: КнигоГрад, 2008. – С. 328–330.
14. Кулагин А.Ю. Сравнительная экология ивы корзиночной и
ивы шерстистопобеговой // Экология. – 1982. – №4. – С. 51–55.
15. Милютин Л.И. Генетико-эволюционные основы устойчивости
лесных экосистем // Лесоведение. – 2003. – №1. – С. 16–20.
16. Правдин Л.Ф. Ива, её культура и использование. – М.:
Изд-во АН СССР, 1952. – 168 с.
17. Скворцов А.К. Ивы СССР. – М.: Наука, 1968. – 262 с.
18. Старова Н.В. Селекция ивовых. – М.: Лесн. пром-сть,
1980. – 206 с.
19. Сукачев В.Н. Работы по селекции ивы // Лесн. хоз-во. –
1939. – № 3. – С. 24-34.
20. Цвелев Н.Н. Определитель сосудистых растений
Северо-Западной России (Ленинградская, Псковская и Новгородская области). –
СПб.: Изд-во СПХФА, 2000. – 781 с.
21. Шабуров В.И. Экспериментальная межвидовая гибридизация
в роде Salix // Третий съезд ВОГиС им. Н.И.Вавилова. – Л.: Наука,
1977. – С. 515.
22. Belyaeva I.
Nomenclature of Salix fragilis L. and
new species, S. euxina (Salicaceae)
// Taxon. – 2009. – 58(4). – P. 1344–1348.
23. Belyaeva I.
Tipification of Pallas′ names in Salix / I. Belyaeva, A. Sennikov
// Kew Bulletin. – 2008. – Vol. 63. – P. 277–287.
24. Bollmark L.
Seasonal dynamics and effects of nitrogen supple rate on nitrogen and
carbohydrate reserves in cutting-derived Salix
viminalis plants / L. Bollmark, L. Sennerby-Forsse, T. Ericsson // Can. J.
Forest Res. – 1999. – 29, № 1. – P. 85–94.
25. Čhmelar J. Die
Weiden Europas / J. Čhmelar, W. Meusel. – Wittenberg–Lutherstadt: 1976. –
143 s.
26. Dawson W.M. Clonal
selection in willow (Salix) grown as short rotation coppice for energy
production / W.M. Dawson, A.R. McCracken // Ann. Appl. Biol. – 1998. – 132,
Suppl. – P. 56–57.
27. Håkansson A.
Chromosome Numbers and Meiosis in certain Salices // Heriditas. – 1955. – B.
41. – № 3–4. – S. 454–482.
28. Rothmaler W.
Exkursionsflora. Kritischer Band. – Berlin, 1976. – 811 s.
29. Wilkinsson J. The
cytology of Salix in relation to its taxonomy // Ann. Bot. NS. – 1944. – 8. –
P. 269–283.
METAGENOFUND OF
SYMPATRIC POPULATIONS WITH VARIOUS LEVELS OF REPRODUCTIVE ISOLATION: ON THE
EXAMPLE OF WILLOWS (SALIX L.)
А.А. Afonin, L.N.
Anishchenko, E.V. Borzdyko, N.V. Markelova
Bryansk State Academician I.G.
Petrovsky University
Abstracts.
Suggested term «metagenofund» to define a set of population genofunds as part
of biocoenosis. In case of high probability of hybridization metagenofund
considered a single genetic system: on the example of ecological-genetic
complex Salix alba – Salix fragilis.
In case of low probability of hybridization metagenofund considered a
conglomeration of population genofunds: on the example of cenosis
Salix viminalis – Salix dasyclados.
Keywords: willows, metagenofund,
genetic safety, polyploidy,
hybridization, reproductive isolation.
Афонин Алексей Алексеевич
Доктор с.-х. наук, профессор Брянского государственного университета
Зав. лабораторией популяционной цитогенетики НИИ ФиПИ БГУ
E-mail: afonin.salix@gmail.com
ИВЫ: Теоретическая и прикладная саликология
ОБЩАЯ И ТЕОРЕТИЧЕСКАЯ БИОЛОГИЯ